Organogensis
VI.1 COORDINATORS
VI.2 PARTICIPANTS
VI.3 SUMMARY
VI.4 INTRODUCTION
VI.5 SPECIFIC AIMS
VI.6 BACKGROUND AND SIGNIFICANCE
- VI.6.i General Background to Organogenesis
- VI.6.ii Interplay of Physical and Genetic Mechanisms in Organogenesis
- VI.6.ii.a Cell Adhesion
- VI.6.ii.b Chemotaxis and Haptotaxis
- VI.6.ii.c Epithelial and Mesenchymal Tissues
- VI.6.ii.d Epithelia and Cell Polarity
- VI.6.ii.e Mesenchyme and Extracellular Matrix
- VI.6.ii.f Mesenchymal Condensation
- VI.6.ii.g Cell Excitability
- VI.6.ii.h Reaction-Diffusion Mechanisms
- VI.6.ii.i Differentiation
- VI.6.ii.j Summary of Basic Mechanisms of Pattern Formation and Morphogenesis
- VI.6.ii.k Neuronal Guidance, Fasciculation and Defasciculation
- VI.6.iii Background to Specific Experimental Systems
VI.8 PRELIMINARY RESULTS
VI.9 RESEARCH DESIGN AND METHODS
VI.10 RELATIONSHIP TO CYTOSKELETON (PROJECT 2) AND BIOLOGICAL NETWORKS (PROJECT 1)
VI.11 TIMELINE
< Previous | Page 4 of 27 | Next >
VI.6.ii.h Reaction-Diffusion Mechanisms:
Reaction-diffusion mechanisms are the continuum simplification of the complex interactions between cells and their local environment. For diffusible chemicals, the biochemical kinetics that cause chemical oscillations can induce standing or traveling spatial periodicities of chemical concentration, e.g., via Turing's "reaction-diffusion" instability (1952). Thus the mechanisms of sequential segmentation in "short germ band" insects such as beetles (Patel et al. 1989), simultaneous segmentation of Drosophilae (Fig. VI.1c), and vertebrate segmental plate generation may have a common evolutionary origin. embryogenesis establishes the body plan, similar processes on a local scale can generate organ primordia and establish their characteristic structures. For example, a Turing-type reaction-diffusion mechanism apparently alters local adhesive properties in chicken skin epithelium (Jiang et al., 1999), giving rise to the epithelial placodes that provide the primordia for developing feathers (Chuong and Edelman, 1985). In fish skin a similar mechanism controls pigment production, inducing colored stripes or spots (Kondo and Asai, 1985). Comparative studies of epithelial development in butterflies and fruit flies have shown that the same set of underlying morphogens can regulate either integumentary color pattern or tissue shape, depending on the species (Carroll et al., 1994).
VI.6.ii.i Differentiation:Apart from their ability to change their adhesive properties, cells can also vary other functions: contractility (muscle), support (cartilage, bone), membrane excitability (neurons), oxygen transport (erythrocytes), etc.. This diversification, which occurs at the level of gene expression, is referred to as differentiation. Differentiation in solid tissues must occur in a well-regulated spatial pattern to ensure appropriate function of the resulting organs.
VI.6.ii.j Summary of Basic Mechanisms of Pattern Formation and Morphogenesis:The diagram below summarizes the preceding discussion and indicates how the fundamental "generic" properties of cell-cell adhesion and differential adhesion, cell excitability, and cell polarity can generate recognizable properties of embryos and developing organs. In many cases these generic mechanisms may be just a starting point both developmentally and evolutionarily. In particular, elaborately fine-tuned genetic mechanisms may have been built on these generic forms during phylogeny and ontogeny to produce the complex developmental mechanisms found in modern organisms (Newman and Müller, 2000).
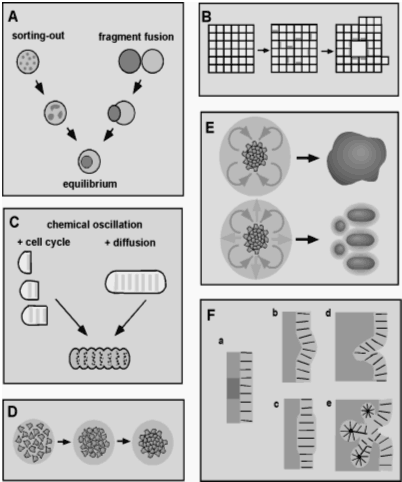
|
| Fig VI.1 (A) Schematic representation of the behavior of intermixed cells and corresponding tissue fragments for two differentially adhesive cell populations (Steinberg, 1998). (B) Schematic view of the formation of a lumen or internal cavity by differential adhesion in an epithelioid tissue consisting of polarized cells (Newman, 1998). (C) Schematic representation of two modes of tissue segmentation that can arise when the tissue's cells contain a biochemical circuit that generates a chemical oscillation or "molecular clock," and the oscillating species directly or indirectly regulates the strength or specificity of cell adhesivity (Newman, 1993). (D) Schematic representation of mesenchymal condensation, which occurs during skeletal morphogenesis and many other developmental processes. (E) Morphogenesis of connective tissue, such as cartilage rods and nodules, occurs by the interplay of diffusible activators and inhibitors of mesenchymal condensation (Newman, 1996; Newman and Tomasek, 1996). (F) Types of epithelial morphogenesis (Newman and Müller, 2000). |
VI.6.ii.k Neuronal Guidance, Fasciculation and Defasciculation:
While innervation of tissues uses many of the same diffusible signal and adhesion mechanisms as other tissue morphogenesis, it also presents a number of differences. Axonal processes of neurons can connect over large distances, raising new topological issues for modeling, which require explicit algorithms in addition to the CPM.
Many aspects of neural innervation are self-organizing (Segev and Ben Jacob, 1998). Innervation involves both chemotaxis (globally) and differential adhesion (locally) (Lance-Jones, 1988).
During nervous system development, neurons send out axons, which migrate to their targets and form neuronal connections. One mechanism which guides migrating axons is the extracellular diffusion of chemoattractants (e.g., netrins, neurotrophins, and neurotransmitters) and chemorepellants (e.g., semaphorins). Molecules in the ECM and on the surface of cells (CAMs, e.g. cadherins) can attract and repel axons by contact attraction and repulsion (Oberlander and Tuan, 1994; Widelitz et al., 1993). In addition, nerve fibers can themselves secret diffusible factors such as neurotransmitters (Young and Poo, 1983; Zheng et al., 1994), dynamically altering the spatiotemporal distribution of diffusible factors during innervation.
Growing axons often form bundles or fascicles (Nornes and Das, 1972; Jessell, 1991; Dodd and Jessell, 1988). Contact attraction mediated by CAMs on the surface of nerve fibers keeps the bundles together. But how do axons come together in the first place? Simulations (Hentschel and Van Ooyen, 1999) have shown that the global pattern of fasciculation depends on the spatiotemporal distribution of diffusible chemoattractants and chemorepellants.
One universal feature of neuronal projection, which is present in situations as different as the eye, ear, skin, and muscle innervation, is the formation and preservation of a somatotopic map. In a somatotopic map, the spatial relation among individual sources of innervation is preserved through a nerve fiber and reconstructed when the nerves spread out (defasciculate) at the target. How does this spatial organization form?